

Photo by D. Sillman
(Click to access an audible version of this blog)
Almost every living organism can detect chemicals in its environment. From our large, complexly structured, vertebrate perspective, we call this chemosensory ability the “sense of smell” (or, more precisely, “olfaction”). The organ used to accomplished olfaction is, of course, the nose. Over the range of living things, though, “noses” come in many different forms!
The “nose” of a bacterium, for example, is a protein located on the surface of the cell. In E. coli, a gram negative (i.e. double plasma membraned) bacterium that is one of the most common organisms used in molecular biology research, there are specific protein receptors for environmental chemicals both on the inner plasma membrane and in the periplasmic space in between the inner and outer plasma membranes. When a chemical binds with one of these proteins a cascade of receptor and enzyme changes involving methylation and phosphorylation of intermediates occurs that alters the behavior of the bacterium. Further, both inhibition or activation of a single receptor system causes adjacent receptor protein systems to similarly react leading to an amplification of the message and response (condon.com/Biotech/Bacteria-sensors).

Amoeba proteus. Photo by N. Hulsmann, Flickr
The ”nose” of the common protozoan, Amoeba, is also a protein on its plasma membrane. When environmental chemicals bind to these receptor proteins a cytoplasmic G-protein signaling system is activated. Phosphorylation and subsequent dissociation of the G-protein components in the Amoeba’s cytoplasm trigger signaling cascades that lead to physiological (and behavioral) changes in the organism (changes that might include movement toward and endocytosis of attractants or movement away from repellants). G-protein systems are also seen in most olfactory receptors throughout both invertebrate and vertebrate animal phyla! (Guetta, Dorian et al. “FYVE-Dependent Endosomal Targeting of an Arrestin-Related Protein in Amoeba.” Ed. Diane Bassham. PLoS ONE 5.12 (2010): e15249. PMC. Web. 15 July 2018).
The “nose” of a jellyfish is made up of specific chemosensory proteins that cover most of its body surface (both on its dome-shaped bell and also on its tentacles). These epidermal receptors are directly exposed to chemicals in the jellyfish’s environment and trigger G-protein changes in the receptor cells that may stimulate directed movement in the jellyfish or the release of protective nematocysts.
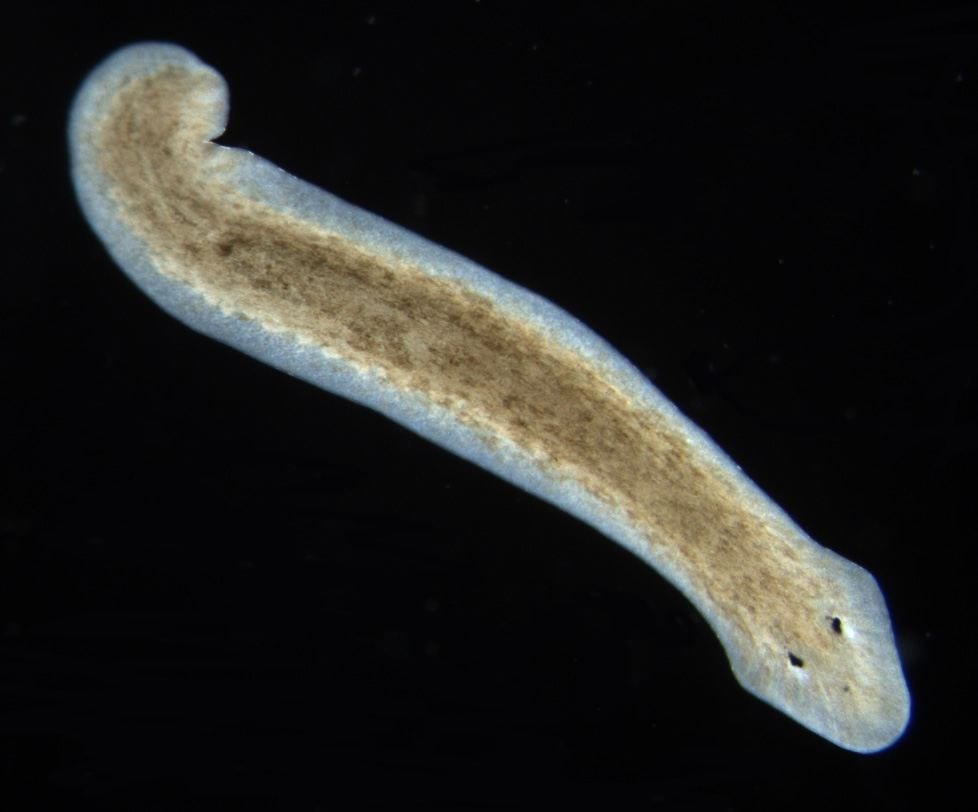
Planaria (Dugesia subtentoculata) . Photo by E. Sola, Wikimedia Commons
Planaria are a type of free-living flatworm. A major evolutionary advance in flatworms over jellyfish is the development of a head! A planarian’s head has two eyespots that are sensitive to light and two lateral projecting auricles (earlike structure) that have both tactile and chemical receptors embedded in them. A flatworm’s “nose”, then, is in its “ears.” (Shirsat N, Siddiqi O. 1993. Olfaction in Invertebrates. Curr Opin
Neurobiol. Aug;3(4):553-7.)
The nematode Caenorhabditis elegans is another extremely common lab organism in molecular biology. Around its anteriorly located lips are sets of sensory structures that house its chemosensory cells (its “nose”). Both volatile and water soluble chemicals can be detected by these cells which then trigger positive responses to food and negative responses to potential dangers or threats. Most of C. elegans’ nervous system and more than five percent of its genes are associated with the recognition of environmental chemicals (there are 600 gene sequences in C. elegans associated with olfactory receptors!) . The G-protein systems in these olfactory receptors is also well documented. (www.wormbook.org/chapters/www_chemosensation/chemosensation.pdf)

Grasshopper. Pixabay.
As invertebrates and then vertebrates develop more and more complex organ systems, the location of the cells that house the plasma membrane bound receptor proteins often becomes more and more specialized. Crayfish “smell” (and also feel) with their antennae as do butterflies and grasshoppers. In fact, these stalked, extended linear “noses” are very common throughout the phylum Arthropoda.
In vertebrates, the chemical receptor proteins (now called olfactory receptors or odorant receptors or “OR’s” for short)) are located on the olfactory sensory neurons in the epithelium in the nose. When an odorant chemical binds to an OR site a G-protein cascade is induced in the olfactory neuron leading to the generation of an action potential (and nerve impulse). Each OR has a range of specific chemicals to which it can react and each odorant chemical has a range of types OR’s to which it can bind. This binding and response complexity makes the olfactory system responsive to an incredibly wide range of odorant chemicals, but causes some ambiguity in the generated sensory information. Novel scents can be readily detected but their precise sensory signaling signature and subsequent identification can be quite obscure.
Photo by A. Levine, Wikimedia Commons
In humans, there are over 900 human OR genes spread out over more than 100 chromosomal locations. Twenty-one of the twenty-three human chromosomes contain OR genes (only chromosome 20 and the Y chromosome lack OR coding). Two-thirds of these human OR genes , though, are non-functional. So somewhere between 300 and 400 human genes are actually coding for OR receptor proteins. Humans have approximately six million OR sites in their olfactory epithelium.
In dogs, our super-smelling best friends, 817 gene sequences for OR’s were identified in a recent study. These genes were found on twenty-four of the dog’s thirty-nine chromosomes in thirty-seven distinct regions. It is estimated that dog’s have about 30% more kinds of olfactory receptors than humans.
The comparison of human and dog OR genes suggests a strongly conserved distribution of genes. This implies that the OR genes of both species (and possibly many other mammals) evolved from a common mammalian ancestral source. These ancestral genes were so useful that they were passed along relatively unchanged through a long succession of evolutionary transformation. The dog OR gene compliment, though, is more expansive and diverse than of humans and contains a rich array of unique, canine OR genes. This suggests that evolution of the dog’s olfactory system involved building on the highly conserved common genes by the addition of novel genetic adaptations.
Dogs in addition to having a greater diversity of OR receptors also has an olfactory epithelial area that is 20 times larger than that of humans. They also have thirty million olfactory receptors (six time more than humans) packed into the olfactory area of their noses. The anatomy of a dog’s nose also funnels a significant percentage of inspired air directly to the olfactory epithelium. Humans must consciously, and vigorously sniff to drive even a small percentage of their inspired air to their olfactory-sensitive cells. Further, the area of a dog’s brain devoted to olfaction is forty times larger than the olfactory area of a human brain. The detection and processing of these environmental scents in a dog is an experience that is probably beyond the comprehension of a human!
One thing that humans can do much better than dogs is drive scent molecules from their pharynxes up into their nasal olfactory epithelium. This type of “smelling” (which is a critical part of tasting!) is called retronasal olfaction (the regular smelling straight in through the nose is called “orthonasal olfaction”). With our human retronasal olfaction we are able to taste the exquisite array of molecular components of our food. Without it, I bet everything would taste like dog food!
(Discussion of OR genes is primarily from Quignon, Pascale et al. “Comparison of the Canine and Human Olfactory Receptor Gene Repertoires.” Genome Biology 4.12 (2003): R80. Print.)
(Coming soon: an olfactory field trip through the woods!)