The integument of the insect thorax is composed of numerous hardened plates (sclerites) that are connected to each other by elastic regions (conjunctivae). Contraction of thoracic muscles move plates along conjunctivae maintaining its functional integrity. Movement of dorsal plate (notum) relative to the lateral one (pleuron) is particularly important in wing bearing thoracic segments; the wing is connected to the notum and pleuron through minuscule sclerites that transmit the movement of thoracic plates into wing flapping.
Separation of hard and rigid thoracic plates by flexible and soft regions certainly increases the vulnerability of the thorax. It is not a big surprise that conjunctivae often disappear during the course of the evolution in brachypterous or wingless insects. “Fusion” of hard plates is perhaps the most distinct in Formicidae where nota of both pterothoracic segments (meso- and metathorax) are fused to the pleura (Fig. 1).

In fully winged Hymenoptera, however, the wing bearing mesonotum and metanotum remained (usually) separated from the rest of the thorax (Fig. 2).

Both nota accommodate a wing pulsatory organ (po2, po3: Fig. 3) that draws out the hemolymph from wing veins through the hollow scutellar arms (sa2, sa3). It is not a big surprise that development of the hymenopteran metanotum is negatively correlated to the size of the hind wing and its venation. In Orthogonalys, for instance, the hind wings are barely reduced comparing to the fore wings and hence are with well developed venation.

Although the metascutellum of fully winged hymenopterans with smaller hind wings (and hind wing venation) are generally reduced in size and even if it is USUALLY well separated from the metapectal propodeal complex. The most reduced metanotum is of Trissevaniini where it is represented laterally by two narrow sclerites (mn3: Fig. 4).

One of the main reasons for using the term USUALLY so often in morphological characterization of Hymenoptera are ceraphronoids.
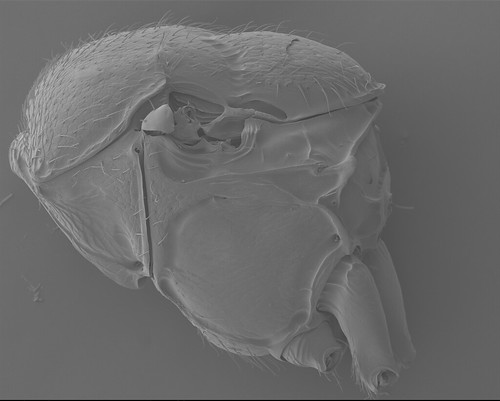
Of course, one of the synapomorphies of the superfamily is the absence of the metanotum.

Several transverse impressions and row of foveae are at the site where the metanotum should be located and neither of them could be precisely defined as the remnant of the boundary between the metanotum and the more posterior thoracic areas (the large, oblique foveolate line crossing dorsolaterally the complex is the antecostal line (al: Fig. 6) that corresponds to the third phragma, an internal ridge that serves of the site of origin of longitudinal muscles connecting the mesosoma to the abdomen). Therefore, we simply don’t know what has happened with the metanotum during the course of the certainly eventful ceraphronoid evolution.
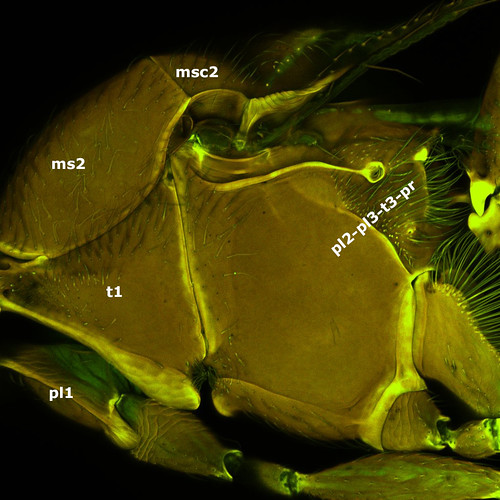
On top of this the mesopectus is fused with the metapectus in this superfamily leaving the mesosoma composed of only 5 sclerites: pronotum (t1), two propleura (pl1), mesoscutum (ms2), mesoscutellum (msc2), and the mesopleuro-metapleuro-metanoto-propodeal complex (pl2-pl3-t3-pr).
Although it is really hard to find an acceptable explanation for the compact mesosoma of Ceraphronoidea, its consequences can readily be observed when we look inside: direct muscles of the fore wings, which normally arise only from the mesopectus, invade the entire pleural surface, while the small direct flight muscles of the hind wing are restricted on a very small region.

What is really easy to do if you work with ceraphs? Writing blog posts about really cool, yet unpublished discoveries! This neglected but really important group of Hymenoptera—don’t forget that ceraphronoids are one of the most abundant Hymenoptera superfamilies—have lots of surprises in store for us … hopefully we will live long enough to capture and expose at least a fraction of them!
Leave a Reply