
 We have shown that 13C direct-detect NMR efficiently generates comprehensive structural data for disordered systems that can be connected with insights into the thermodynamics of their interactions and the mechanisms whereby structure and function are modulated by post-translational modification. Our efforts to describe IDP ensembles, and the ways in which post-translational modification alters them, is already advancing understanding of their diverse biological functions.
We have shown that 13C direct-detect NMR efficiently generates comprehensive structural data for disordered systems that can be connected with insights into the thermodynamics of their interactions and the mechanisms whereby structure and function are modulated by post-translational modification. Our efforts to describe IDP ensembles, and the ways in which post-translational modification alters them, is already advancing understanding of their diverse biological functions.
This has been a continuous project over the past 10 years, highlights of which are reviewed below. We are always on the lookout for prospective postdoctoral associates with a background in NMR spectroscopy who would like to develop pulse programming skills. If you are interested in this opportunity, please email Dr. Showalter.
All NMR pulse sequences discussed on this page (and many more) are available for download in Bruker format through our NMR Pulse Program Library.
Featured Publications
Gibbs, EB, Cook, EC, Showalter, SA. Application of NMR to Studies of Intrinsically Disordered Proteins. Arch Biochem Biophys. 2017; 628: 57-70. PMID: 28502465
Gibbs, EB, Showalter, SA. Quantitative Biophysical Characterization of Intrinsically Disordered Proteins. Biochemistry. 2015; 54: 1314-1326. PubMed PMID: 25631161
Current Directions: Broaden the diversity of IDPs and their complexes amenable to direct structure analysis.
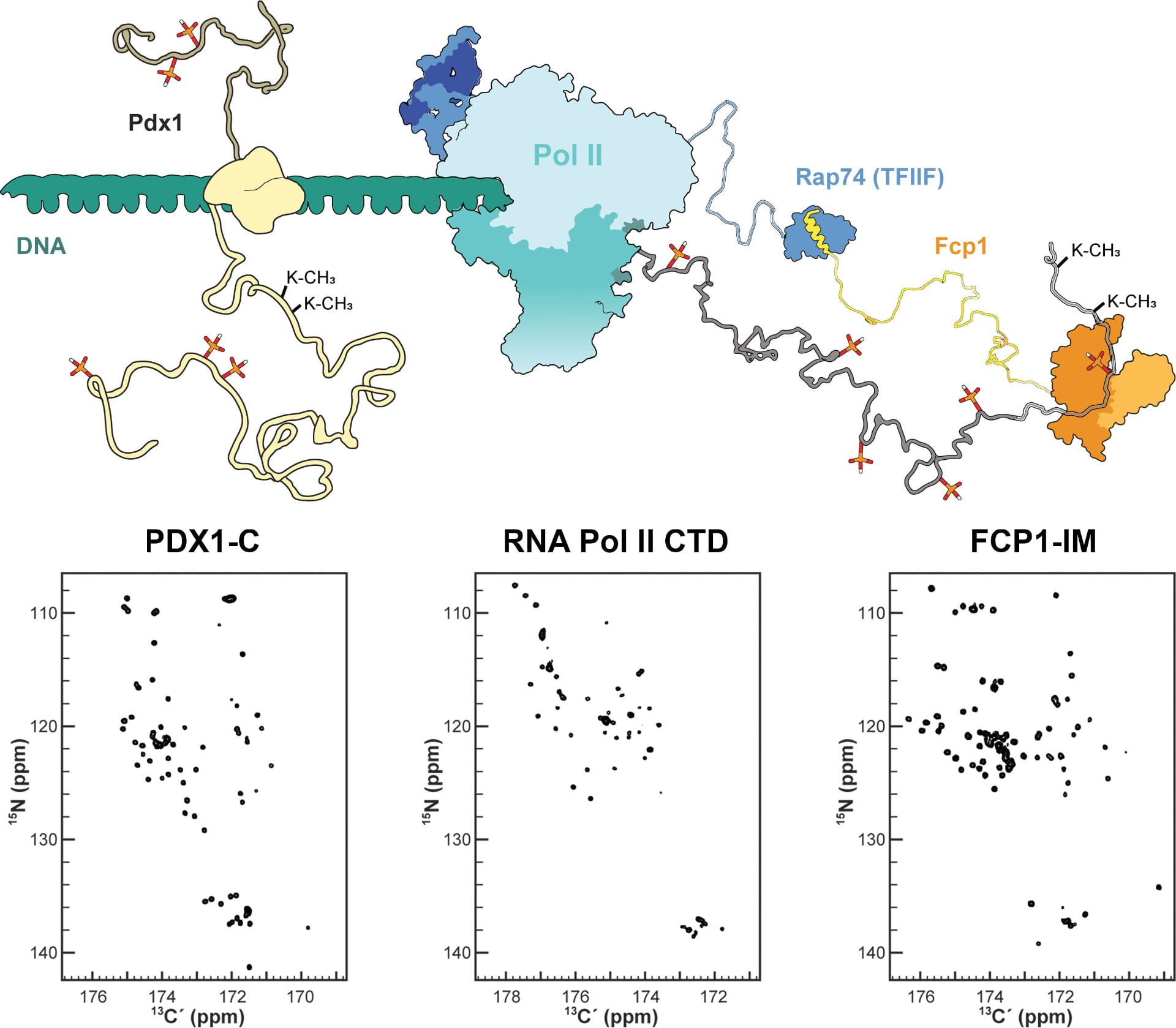
The extreme sequence bias displayed by IDPs presents a major technical barrier that still restricts which proteins can be studied for their binding properties. Through our prior work developing 13C direct-detect NMR, we have extended the range of accessible proteins to include proline-rich IDPs. To further broaden our approach, we now seek to target aromatic-rich IDPs, as exemplified by the RNA Polymerase II C-terminal Domain. While aromatic residues are rare in IDPs, they are over-represented in IDP binding surfaces. To improve characterization of complexes, we are working to develop 13C direct-detect paramagnetic relaxation enhancement in trans and chemical exchange saturation transfer.
Chemical Shift Assignment Through 13C Direct-Detect NMR.
Carbon-detected spectra generally offer superior resonance dispersion and are conducive to the assignment and study of proline-rich constructs. In the case of Pdx1-C, the 1H,15N-HSQC in the accompanying figure demonstrates acceptable resolution, in the sense that most resonances are resolved (nearly) to baseline, but their occupancy of a very narrow chemical shift range is still evident. The contrast between the 1H,15N-HSQC and the 13C,15N-CON experiment shown to the left is dramatic. Pdx1-C features 18 proline residues in its sequence, which are absent in the 1H, 15N-HSQC, but readily identified in the 13C,15N-CON by their characteristic 15N chemical shift between 132-144 ppm.

Most trained biomolecular NMR spectroscopists are familiar collecting a “fingerprint” 2D and then proceeding to triple-resonance assignment strategies where backbone connectivity is generally built through the 13Cα and 13Cβ nuclei. However, aliphatic carbon chemical shifts of IDPs generally do not deviate much from random coil values, which can produce assignment complications for repetitive sequences. Instead, we find great advantage in 3D spectra built from the (CA)CON and (CA)NCO that correlate the backbone amide 15N chemical shifts of pairs or triplets of amino acids. The (HACA)N(CA)CON records the 15N of residue i as an autocorrelation peak with a cross-correlation to the 15N-chemical shift for residue i-1. The (HACA)N(CA)NCO provides complementary information: the indirect 15N-dimension records cross-correlation peaks to the 15N-resonance of residues i-1 and i+1. Use of these experiments is illustrated in the figure above through the 3D strips on the right, which represent residues 255-259 from Pdx1-C. The corresponding resonances in the 2D 13C,15N-CON are colored in red as a visual reference.
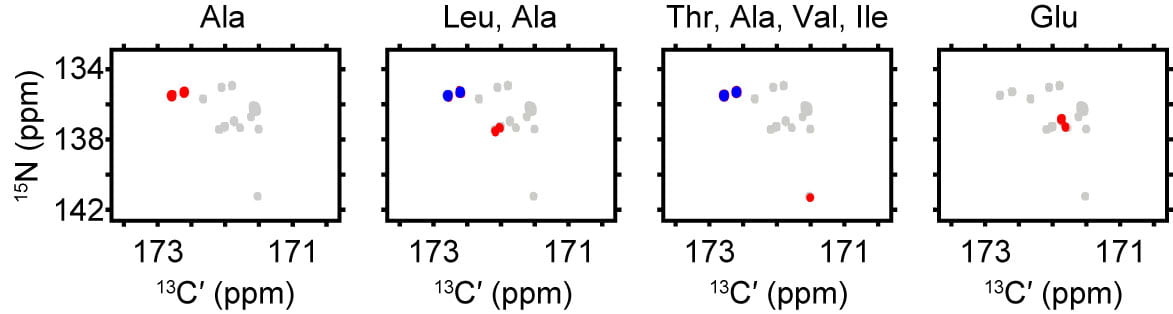
In addition, carbon-detect amino acid-specific (CAS NMR) variants of the 13C,15N-CON spectrum provide a powerful complement to 3D-based assignments. The utility of this technique is especially clear in the context of the resonances that arise from peptide bonds where proline contributes the 15N-atom. In Figure 1c, we see how to combine the A-, TAVI-, LA-, and E-specific spectra to accelerate assignment of the proline resonances, which is advantageous for a proline-rich system like Pdx1-C.
Featured Publications
Cook, EC, Usher, GA, Showalter, SA. The use of 13C Direct-Detect NMR to Characterize Flexible and Disordered Proteins. Methods Enzymol. 2018; 611: 81-100. PMID: 30471706
Bastidas, M, Gibbs, EB, Sahu, D, Showalter, SA. A Primer for Carbon-Detected NMR Applications to Intrinsically Disordered Proteins in Solution. Con Mag Reson A. 2015; 44A: 54-66. PMID: In process.
Sahu, D, Bastidas, M, Showalter, SA Generating NMR Chemical Shift Assignments of Intrinsically Disordered Proteins Using Carbon-Detect NMR Methods. Anal Biochem. 2014; 449: 17-25. PMCID: PMC3944900
Structural and Dynamic Constraints from 13C Direct-Detect NMR.
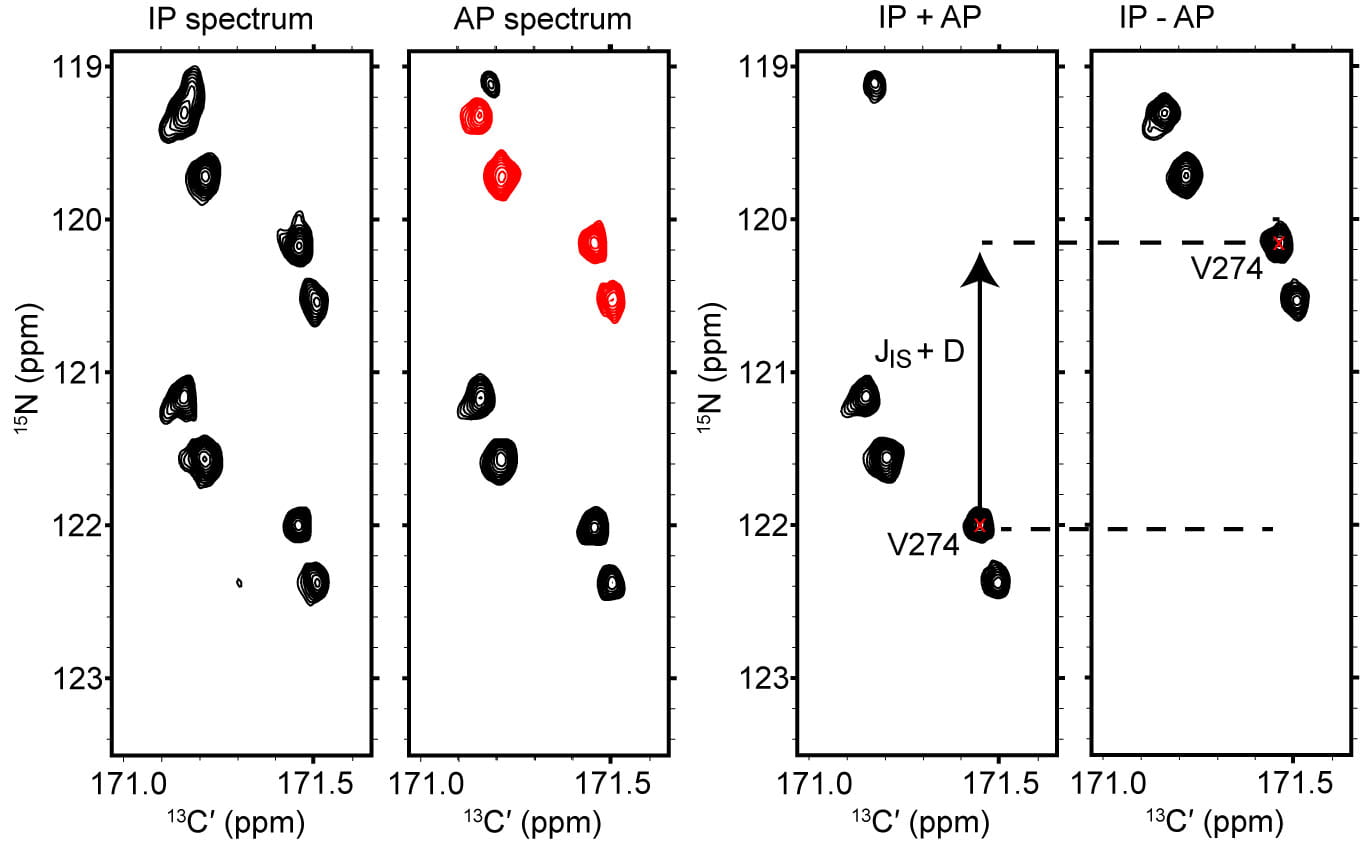
As a supplement to existing 1H-detected RDC experiments, we have developed 13C,15N-CON variants that encode the NH, C´Cα, C´N, and HαCα RDC. In all cases, RDCs are recorded through a pair of spectra collected using the “in-phase (IP) / antiphase (AP)” technique in which the scalar coupling between the two nuclei under investigation is not refocused during the indirect chemical shift evolution period, resulting in the pair of spectra like those depicted on the left in the figure below. In the IP spectrum, both components of the doublet are phased equivalently, whereas the AP doublet features a 180° phase offset between its two components. Addition and subtraction of the IP and AP spectra leads to the spectra depicted on the right in the figure below, with the advantage that the scalar coupling constant can be determined from the peak offset without adding spectral crowding. Comparison of the splitting in the isotropic and aligned sample yields the RDC value for each spin pair through subtraction.
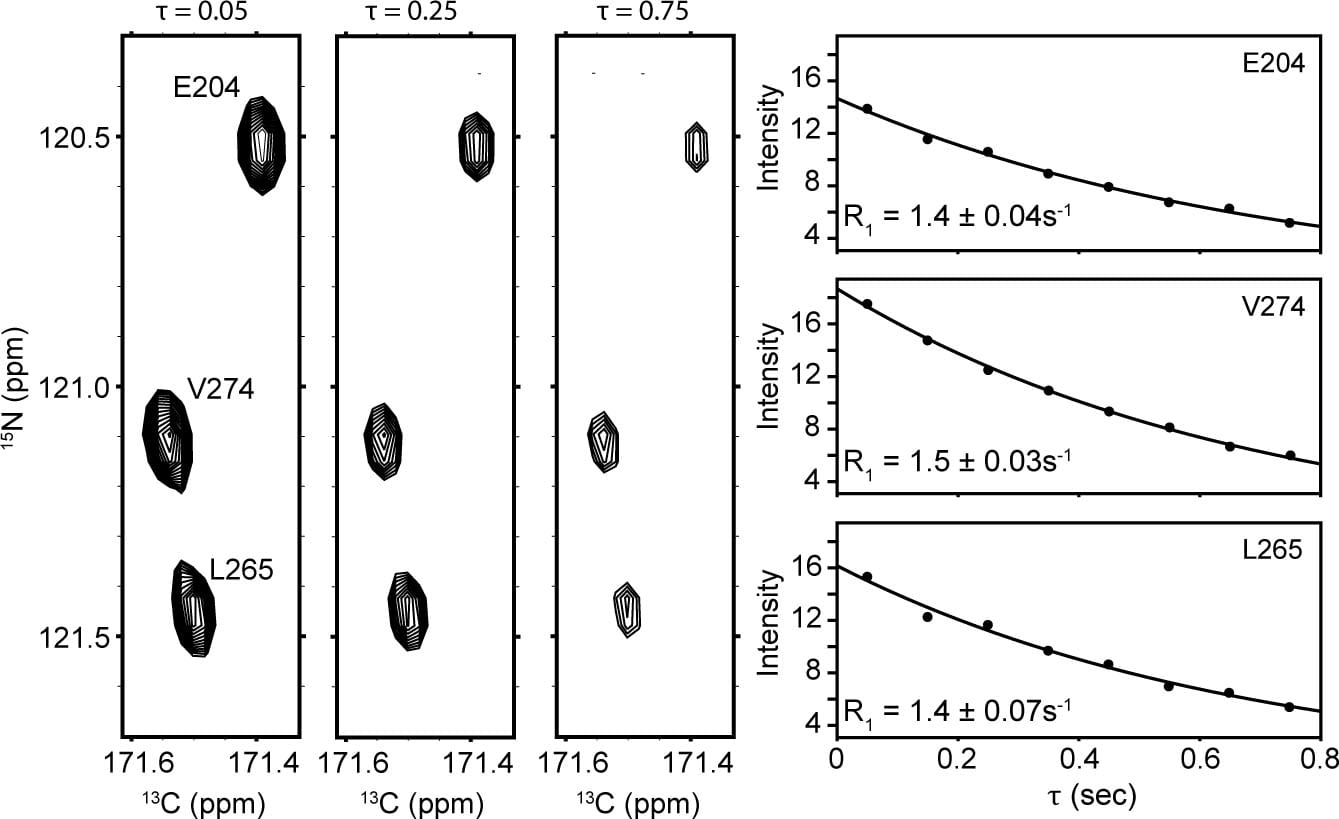
Measurement of 15N spin relaxation is a routine method used to assess protein backbone flexibility in both well-folded and intrinsically disordered systems. In 2012, we reported adaptation of the 13C,15N-CON to encode 15N R1 and R2 relaxation rates. Collection in a 13C direct-detect format is directly analogous to standard 1H-detected experiments. For example, residues E204, V274, and L265 from Pdx1-C display mono-exponential relaxation, as demonstrated below.
Featured Publications
Cook, EC, Sahu, D, Bastidas, M, Showalter, SA. The Solution Ensemble of the C-Terminal Domain from the Transcription Factor Pdx1 Resembles an Excluded Volume Polymer. J. Phys. Chem. B. 2019; 123: 106-116. PMID: 30525611
Lawrence, CW, Showalter, SA. Carbon-Detected 15N NMR Spin Relaxation of an Intrinsically Disordered Protein: FCP1 Dynamics Unbound and in Complex with RAP74. J Phys Chem Lett. 2012; 3: 1409-1413. PubMed PMID: 26286791
Funding Acknowledgement
![]()
We are deeply grateful to the National Science Foundation for their sustained support of this research program through the following grants:
MCB-1932730
MCB-1515974
MCB-0953918